
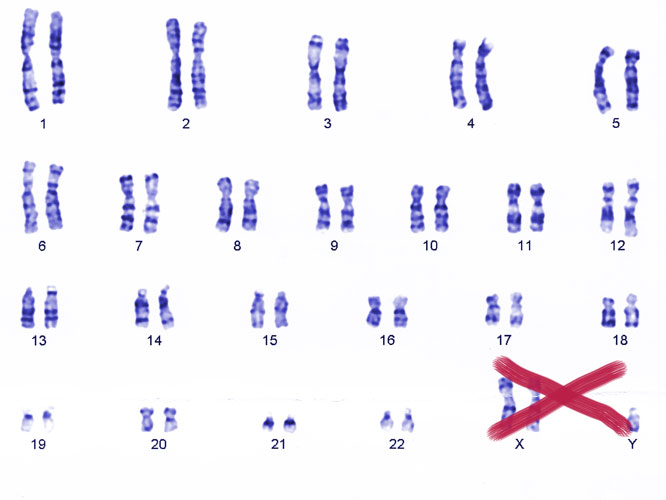
One of the major risk factors for autoimmune diseases is being born with two copies of the X chromosome. For example, women—who typically carry two Xs—face around ten times the risk of lupus, while men with lupus are around 15 times more likely than the general population to carry two Xs and a Y (Klinefelter syndrome), rather than the usual single X and Y.
The luck of the sex-chromosome draw affects many other complex diseases in prevalence, severity, symptoms, and rates of progression. Women face about twice the risk of depression, for example, while men are three to five times more prone to liver cancer. Though environmental factors like sex hormones and alcohol consumption are behind some of the differences, Alon Keinan (Cornell University) argues that genetic variation carried on sex chromosomes must also play a role.
But how big is that role? He says there is little conclusive evidence. The problem is, the sex chromosomes have been largely ignored in the flood of human genome-wide association studies (GWAS) of recent years.
“We are conducting all these genome-wide studies, but sometimes forget that the genome also includes sex chromosomes,” he says. There are many other unknowns in how sex chromosomes affect complex disease, including the population-wide variation in X-chromosome inactivation, a crucial factor in the expression of X-linked variants.
For the 2014 American Society for Human Genetics meeting in October, Keinan teamed up with Melissa Wilson Sayres (Arizona State University) to organize a session called “The X-Factor of Complex Disease: From Evolution to Association Studies of the X Chromosome” to discuss these issues and more. The idea was to bring together speakers with expertise ranging from evolutionary to functional genetics to present research on sex chromosomes and their relevance to interpretations of human medical genetic data.
“Part of the motivation is that medicine today is often driven by diagnosis and treatment of males. We would like to better understand the genetic risk factors that affect males and females differentially, to make at least one step towards sex-specific diagnosis and treatment,” says Keinan.
GSA Journals Assistant Editor Cristy Gelling caught up with Wilson Sayres and Keinan to discuss X-linked disease associations and the evolution of X-inactivation.
Bringing X back
Although the X chromosome makes up about 5% of the human genome and carries 4% of all genes, it is linked to fewer than 0.5% of associations in NHGRI’s GWAS catalog. For example, chromosome 7, similar in size to the X, has about 10 times as many associations. In model organisms, sex chromosomes are responsible for more phenotypic variation than you would predict from their size. But however you do the math, we’ve likely missed the vast majority of the X-linked associations with complex disease.
The main problem is that most GWAS discard all data from the X chromosome. “It’s understandable,” says Keinan. “You can imagine when researchers are competing to publish quickly, it’s easy to ignore something that is only 5% of the genome and needs extra time and special expertise.”
Even when the X is included, it’s typically analyzed using methods designed for autosomes. But standard statistical models for detecting significant associations don’t perform well for X chromosomes because they have unique patterns of diversity. For example, because recombination is suppressed in sex chromosomes, they evolve very differently from autosomes. One speaker at the session, Krishna R. Veeramah of Stony Brook University, compared the effects of natural selection on the X chromosomes to the autosomes in humans and apes. The X chromosomes in most of these species (including humans) show evidence for greater purifying selection and faster adaptive evolution rates compared to autosomes.
Sex-biased social practices, such as polygyny and male migration also affect variation on the X chromosome in ways that can distort association tests. One particularly recent example of a sex-biased demographic effect was caused by human slavery in the United States. Today’s African-American populations carry, on average, a higher proportion of African ancestry on their X chromosomes compared to autosomes. This shows much of the European genetic heritage of African-Americans is derived from male, rather than female ancestors—likely the legacy of white men fathering children with the women they enslaved.
It’s not only the different models of variation that create headaches when analyzing X-linked GWAS data. All the other steps of analysis, including determining an individual’s genotype, filtering out poor quality data, and imputation of missing genotype values, can be distorted by the difference in copy number between sexes. The end result of applying autosomal pipelines to the X chromosome is lower statistical power and a higher rate of false positives.
In his presentation, Keinan described a new “XWAS” analysis pipeline developed by several graduate students and postdoctoral fellows in his lab tailored to account for the biology of X chromosomes. To test the performance of the pipeline, the group used it to reanalyze existing autoimmune disease GWAS data for X-linked associations. The results replicated several previously found associations and revealed new candidate disease risk genes on the X chromosome with plausible links to immune function.
So can we now reanalyze the back-catalog of previously performed GWAS and unearth all those missing X-linked associations? Not quite. Although many GWAS datasets are available for reanalysis, they are usually deposited as processed data that have already been through quality control steps. In most cases, these QC steps have not been corrected for sex, and in some cases, the X chromosome data have even been discarded. Keinan hopes more geneticists will start depositing their GWAS results as raw data.
“All I would like to be able to do at this point is take a look at data that’s already available. A huge amount of taxpayer money has been invested in GWAS, which has paid off in many discoveries and can pay off further if the data generated is available for reanalysis,” he says.
Escape from X-inactivation
Another major complication for modelling how an X-linked variant contributes to disease is X inactivation, a process in which one of the two X-chromosomes in genetic females is randomly silenced. Which of the two chromosomes gets silenced varies from tissue to tissue and cell to cell. That means a particular X-linked variant may or may not be expressed in the tissues relevant to the disease.
X-inactivation doesn’t affect the entire chromosome; around 15 percent of genes “escape” inactivation and are expressed. Adding to this complexity, around ten percent of the genes that escape silencing only do so in some individuals and not in others. At the last talk of the session, Christine Disteche (University of Washington) presented results suggesting the genes that escape X inactivation also vary between tissues, resulting in sex-specific differences in phenotypes. But taking these phenomena into account in models of disease association is not possible because we simply don’t know enough about X-inactivation and its variation in humans, says Wilson Sayres.
“Right now, what we know about X-inactivation heterogeneity is based on nine cell lines, and we don’t even know which populations the lines came from,” she says. She argues that assaying patterns of X-inactivation across populations and within individuals is one of the challenges the field needs to address before it can fully understand disease phenotype variation.
Interestingly, understanding the evolution of sex chromosomes may shed some light on patterns of variation in X inactivation. One of the topics Wilson Sayres discussed at the meeting was how gene loss on the Y chromosome influences escape from silencing on the X.
In the evolutionary past, the X and Y chromosomes were homologous, carrying mostly the same set of genes. But over time, the Y has lost many genes, leaving it much smaller than the X. Wilson Sayres set out to test the idea that X inactivation of a gene only evolves when the gene’s homolog on the Y (technically a “gametolog”) is lost. Silencing one copy of the gene in females would balance out the mismatch between the sexes.
If this is the case, genes on the X chromosome were once expressed from both copies in females (i.e. they “escaped” silencing), but only became subject to X inactivation when their Y gametolog lost function. By re-examining the X inactivation data from the nine cell lines derived from different people, Wilson Sayres and colleagues confirmed that X linked genes with a Y gametolog escape silencing. In contrast, genes with no gametolog on the Y were, on average, silenced in most cell lines.
Crucially, genes with non-functional gametologs—which are earlier along in the process of being lost from the Y—escaped silencing at a greater rate than those that had completely lost the Y gametolog. That suggests an evolutionary lag between loss of the gene on the Y, and the onset of silencing of the gametolog on the X. This model predicts that the genes that still “escape” silencing even though their Y gametolog is no longer function, will eventually be silenced. “This process is still happening in us,” says Wilson Sayres. “There’s ongoing evolution.”
One implication of this continuing adjustment is that different species are at different stages in the evolution of X inactivation. The process is more tightly regulated in mice, perhaps because the mouse Y chromosome has lost comparatively more of its genes than the human Y. A much smaller proportion of genes escape silencing, and there may be lower variation between individuals.
“It might actually be less messy in mice than in humans,” says Wilson Sayres. This has implications for mouse models of human disease, particularly in cases where the phenotype is modified by genes on the X chromosome.
Solve for X
There is much work to do, Keinan and Wilson Sayres agree. Keinan says researchers need to re-analyze the 2000 or so available GWAS, not to mention the newer sequence-based datasets, most of which still do not consider X. He wonders what proportion of the infamous “missing heritability” of complex human traits might be explained by X-linked variants, but more importantly, hopes we can pin down specific variants and pathways that contribute to the sexual dimorphism of human disease.
Wilson Sayres argues that we need to survey human variation in X-inactivation not only across populations, but within individuals, to get a handle on the patterns of tissue mosaicism in X silencing. She would also like to see more diversity incorporated into association studies. One example would be to include individuals with sex chromosome aneuploidies, instead of excluding them from analysis.
“In Turner Syndrome — that’s where you have only a single X — there’s huge variation in phenotype. Some people don’t even know they have it until they do a cheek swab,” she says. “A huge challenge in studying sex-chromosome aneuploidies is that we don’t understand X variation in karyotypically-common people, let alone for those with single or multiple copies.”
But that’s for the long term, she says. “The first step is just to include the X!”
Cristy Gelling is a science writer, lapsed yeast geneticist, and former Communications Director at the GSA.
View all posts by Cristy Gelling »Read more in
-

Thank you, GSA community!
Thank you for being a member of the Genetics Society of America! As GSA’s current president, I am writing to tell you about Society projects and initiatives that we hope you will find useful in advancing your science and your career. Scientific research is a collaborative and exciting endeavor. Scientific societies like GSA exist to…
-

Where are they now? Rosalind Franklin Young Investigator Award recipients share updates on their research
Rosalind Franklin Young Investigator Award applications are open–make sure you submit your application or nomination of a colleague by September 30, 2024.
-

University of Minnesota researchers map genome of the last living wild horse species
The study, published in G3: Genes|Genomes|Genetics, is part of larger conservation efforts to save Przewalski’s horse.
-

Congratulations to the Spring 2024 DeLill Nasser Awardees!
GSA is pleased to announce the recipients of the DeLill Nasser Award for Professional Development in Genetics for Spring 2024! Given twice a year to graduate students and postdoctoral researchers, DeLill Nasser Awards support attendance at meetings and laboratory courses. The award is named in honor of DeLill Nasser, a long-time GSA supporter and National Science Foundation…
-

Carolyn Damilola: an NFS Rising Scientist on a lifelong quest to learn more
Carolyn Damilola is an NFS Rising Scientist from Nigeria doing respiratory system research and paving the way for scientists from underrepresented communities through mentorship.
-

What does a good microgrant proposal look like?
Members of the Microgrant Review Committee share their tips for a successful proposal.
-

The first piece of the facial recognition puzzle
New research in GENETICS gives a first peek at the molecular pathway involved in recognizing faces.
-

New Senior Editor Amy MacQueen joins GENETICS
A new senior editor is joining GENETICS in the Genome Integrity and Transmission section. We’re excited to welcome Amy MacQueen to the editorial team.
-

Block party on the zebrafish sex chromosome
Research in G3 identifies a gene regulatory block of the zebrafish genome responsible for overseeing the maternal-to-zygotic-transition.
-

Unraveling the mysteries of duckweed: epigenetic insights from Spirodela polyrhiza
Research published in G3 offers insight into the impact of DNA methylation on clonal propagation in asexually reproducing plants.
-

A microbiologist’s quest to understand CRISPR in bacterial self-defense
2024 Genetics Society of America Medal recipient Luciano Marraffini determined how CRISPR-Cas systems destroy genetic targets with precision, paving the way for gene editing technology development.
-

Unlocking mysteries of trait and disease heritability in dogs
2024 Edward Novitski Prize recipient Elaine Ostrander, a pioneer of the domestic dog model, discovered numerous genes affecting dog size, morphology, behavior, and disease susceptibility—many of which have relevance in humans.
-

GSA and collaborators Personal Genetics Education & Dialogue and Reclaiming STEM Institute launch NSF-funded BIO-LEAPS project to support culture change in genetics
We are thrilled to announce that the Genetics Society of America (GSA) is collaborating with the Personal Genetics Education & Dialogue (PGED) based in the Department of Genetics at Harvard Medical School, and the Reclaiming STEM Institute (RSI) on a Leading Culture Change Through Professional Societies of Biology (BIO-LEAPS) grant from the U.S. National Science…
-

Daman Saluja: Navigating Science and Policy in India
In the Paths to Science Policy series, we talk to individuals who have a passion for science policy and are active in advocacy through their various roles and careers. The series aims to inform and guide early career scientists interested in science policy. This series is brought to you by the GSA Early Career Scientist…
-

A fly geneticist’s journey into discovering rules of organ development
2024 George W. Beadle Award recipient Deborah Andrew discovered new genes and pathways in Drosophila salivary gland organogenesis. Now, her work can help optimize cell secretion in therapeutic applications and fight malaria.
-

Małgorzata Gazda: How receiving the DeLill Nasser Award helped her land her dream job
Have you ever experienced an event that changes the course of your life, or in this case, your career? Małgorzata (Gosia) Gazda is Assistant Professor at the University of Montreal and in 2022, she received the DeLill Nasser Award for Professional Development in Genetics, which she used to attend and present at the 2022 Population,…
-

Hongyu Zhao joins GENETICS as new Senior Editor
A new senior editor is joining GENETICS in the Statistical Genetics and Genomics section. We’re excited to welcome Hongyu Zhao to the editorial team.
-

GSA Member Julio Molina Pineda Receives DeLill Nasser Award, Shines at TAGC 2024
“At any career stage, the GSA membership is an amazing investment for any genetics professional!” Julio Molina Pineda is a PhD Candidate in Cell and Molecular Biology and a Research Assistant at the University of Arkansas, and a Doctoral Academy Fellow at the Lewis Lab. In 2023, Julio was awarded the DeLill Nasser Award for…
-

In Memoriam: Ellsworth Herman Grell (1932–2023), a pioneer of Drosophila genome engineering and annotation
Ellsworth (Ed) Grell blessed the Drosophila community through three enduring legacies: as a pioneer of chromosome mechanics, as a primary organizer and synthesizer of genetic knowledge in Drosophila, and as a graceful mentor to those fortunate to have known him personally. Ed grew up in rural Nebraska, completed his undergraduate studies at Iowa State, and…
-

Congratulations to the #Fungal24 Poster Award winners!
We are pleased to announce the recipients of the GSA Poster Awards for posters presented at the 32nd Fungal Genetics Conference! Undergraduate and graduate student members of GSA were eligible for the awards, and a hard-working team of judges made the determinations. Congratulations to all! Felicia Ebot Ojong, The University of Georgia My research is focused…
-

Poster presentation tips for TAGC 2024
You’ve been selected to present a poster at The Allied Genetics Conference 2024 in March—you’ve celebrated, made plans to attend, now what? This is an exciting opportunity to showcase your research and engage with fellow members of the genetics community, so you want to make sure you’re prepared. We wanted to offer you some tips…
-

Maximize your TAGC 2024 experience
A guide to all that National Harbor & DC have to offer Are you joining us for The Allied Genetics Conference 2024 in March? Make the most of your #TAGC24 experience in National Harbor! We know the science will keep you busy, but you deserve to unwind and have some fun, so we’ve curated a…
-

Early Career Leadership Spotlight: Sarah Petrosky
We’re taking time to get to know the members of the GSA’s Early Career Scientist Committees. Join us to learn more about our early career scientist advocates. Sarah PetroskyMultimedia SubcommitteeUniversity of Pittsburgh Research Interest I am interested in understanding adaptation that has been happening recently in populations by dissecting the ways that genes underlying an adaptation…
-

TAGC 2024 Early Career Award Winners
GSA is pleased to announce the winners of the early career awards presented at The Allied Genetics Conference 2024. These awards are specific to particular TAGC communities and recognize early career scientists’ outstanding work on their respective research organisms. The awardees will present their talks in keynote sessions at TAGC 2024. Don’t miss the opportunity…
-

Preeminent geneticists recognized with revamped GSA Awards
In 2022, GSA’s Board of Directors launched an audit to review the five major awards conferred by the Society. Today, we are thrilled to announce the recipients of the reimagined GSA Awards, including the new Genetics Society of America Early Career Medal. The scientists honored this year are recognized by their peers for their outstanding…
-

Fly Board funds outreach programs to spread the word about Drosophila research
In 2020, the Fly Board voted to use part of its reserve fund to support efforts to increase trainee participation as well as equity and diversity in the Drosophila community. An awards committee decides how the money will be spent each year, and from 2020–2022, the committee posted a very broad call for applications from…
-

New members of the GSA Board of Directors: 2024–2026
We are pleased to announce the election of four new leaders to the GSA Board of Directors: 2024 Vice President/2025 President Brenda Andrews Professor, University of Toronto It’s an honor to continue my association with the Society by serving as Vice President of the Board of Directors. I have broad knowledge of the ongoing activities…
-

Policy changes are closing the door on the “American Dream”
The American Dream once promised that talent and hard work could open doors. For early career scientists, those doors are now closing. Research is being paused or studies outright canceled, funding delayed or completely pulled, and careers cut short—not because of bad science, but because of changing political priorities. Researchers are being forced to choose: leave science or leave the country. These setbacks send a clear message: the U.S. commitment to supporting the next generation of scientists is waning.
-

Congratulations to the Worm 2025 GSA Poster Award Recipients
We are pleased to announce the GSA Poster Award recipients from the 25th International Worm Meeting! Undergraduate and graduate student GSA members were eligible for these awards, and a hard-working team of judges made the determinations. Congratulations to all! Kehinde Joan Abayomi, Mount Desert Island Biological Laboratory Category: Gene Regulation & Genomics Poster title: “Transcriptome…
-

GENETICS Expands Primers Section
Beth De Stasio of Lawrence University will serve as Senior Editor and work with a newly formed team of six Associate Editors to expand the publication of Primers in GENETICS.